
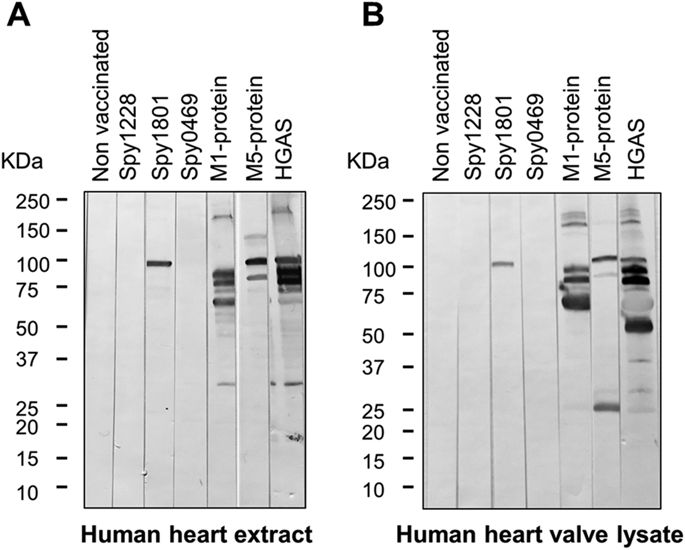
Anti-serum from mice vaccinated with the proteins indicated in the figure was used to test for cross-reactivity towards (A) human heart extract and (B) human heart aorta valve lysate by western blotting. The molecular weight markers in kilo Dalton (kDa) are indicated.
Discussion
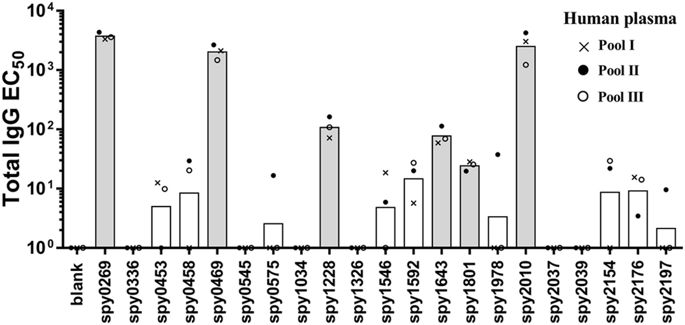
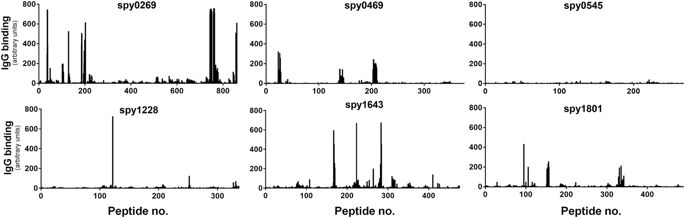
This study describes the identification of three conserved antigens with protective capacity in an experimental GAS infection model. Various strategies have been employed during the last decade to identify protective non-M protein antigens from GAS22,23,24,25,26. Each of these studies identified one to nine protective antigens. Different technical approaches were adopted in these studies, which might explain why only a few antigens were identified in more than one study. Three such antigens were SpyCEP, ScpA and spy0269. Spy0269 was also shown in our study to be recognized by IgG in human plasma (Figs 1 and 2) and by our peptide array to contain several B cell epitopes (Fig. 3).
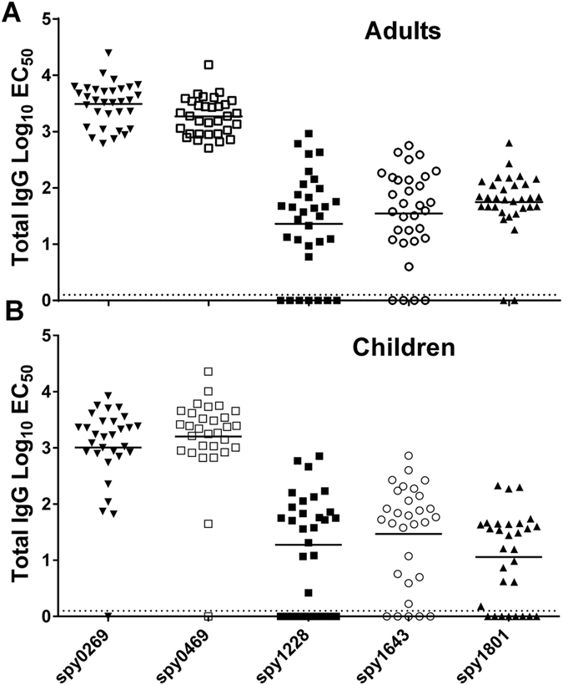
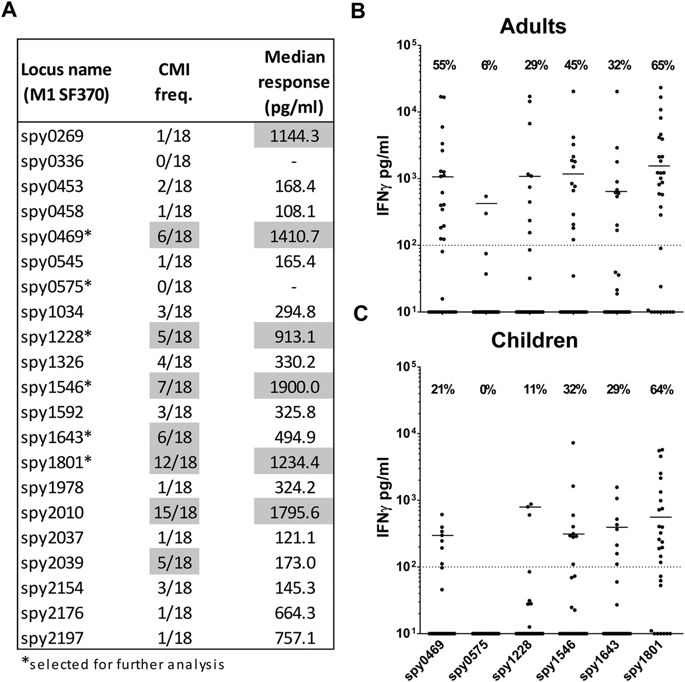
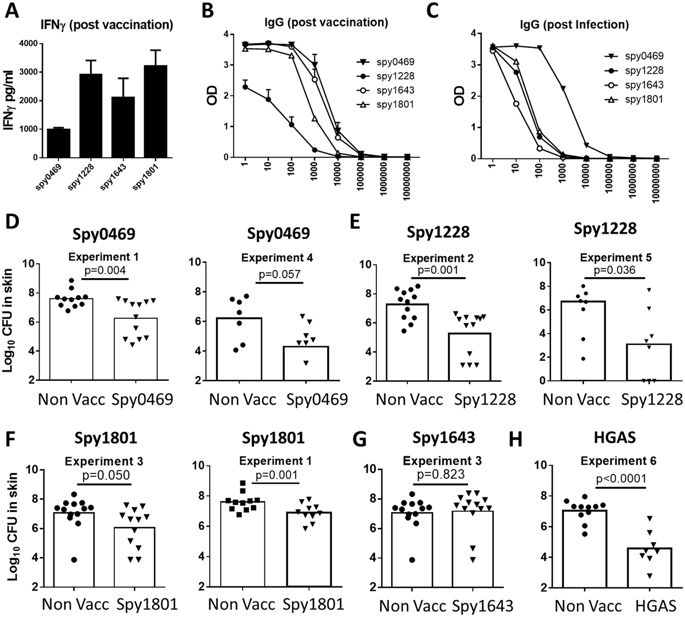
21 antigens passed our initial selection criteria. Six of these were recognized by cellular responses (spy0469, spy1228, spy1546, spy1643, spy1801 and spy2010) and six by antibodies (spy0269, spy0469, spy1228, spy1643, spy1801 and spy2010). Thus, there is a significant overlap between the targets of cellular and antibody responses. The same antigens were also recognized in infected mice suggesting that immunogenicity was independent of host species, and probably related to their expression and availability to the immune system. All individuals, except one child, exhibited adaptive immune responses to at least one GAS antigen, confirming that exposure to this pathogen is very common. In the i.d. infection model, immunization with spy0469, spy1228, and spy1801, but not spy1643, induced a significant reduction in the GAS colonization of the skin (Fig. 5D–G). This is, to our knowledge, the first time these antigens are described as protective antigens.
Spy1228 is a putative conserved lipoprotein35 with unknown function that has previously been shown to elicit an antibody response in mice and humans22. Extracellular location of this protein has also been verified experimentally36. Spy0469 showed strong antibody recognition in all adults and 97% children in our study as well as cellular responses in 55% of the adult donors compared to 21% of the children (p = 0.0198). The biological function of spy0469 is unknown but it contains a LysM domain that is used to locate proteins to the bacterial envelope via non-covalent linkage to peptidoglycan37. The extracellular location of this protein has also been verified experimentally36,38. Spy1801 does not have a confirmed biological function either, but it has been suggested to play a role in biofilm formation as its gene is highly upregulated during biofilm formation39. Consistent with that, computational comparisons indicate that spy1801 belong to a family of peptidoglycan hydrolases that includes a member in Lactococcus lactis that generates peptidoglycan disruptions that are important for biofilm formation40. Spy1801 (that is also annotated as “immunogenic secreted protein” (Isp)), is also known to induce antibody responses in humans41, so it was surprising that anti-sera raised against spy1801 cross-reacted with a 100 kDa protein in two different tissue extracts from the human heart. Although we cannot conclude that the epitope recognized by spy1801 anti-sera in the western blot is available for antibody recognition in vivo and thus play a potential role in GAS pathogenesis, it does raise a safety concern that should be further investigated.
The present work is the first to use a peptide array technology platform to identify linear B cell epitopes in the validation of GAS antibody targets. Using pooled serum from six donors, we confirmed the antigenicity of spy0269, spy0469, spy1228, spy1643 and spy1801 by identification of at least one linear antibody epitope. Subunit vaccines have an upper limit as to how many antigens that can be combined before recombinant expression of the final fusion protein become negatively affected by the molecular weight. However, numerous short linear epitopes like the ones identified in our study could potentially be included in a vaccine construct without obstructing recombinant expression due to size issues. This would allow the vaccine to target a large number of different GAS antigens. Given the high capacity of the array technology, all putative proteins in the GAS genome could be included in a single IgG epitope screen. By that, we were allowed to compare the signal strength of the epitopes from our selected targets with epitopes from every other possible protein in the M1 SF370 GAS genome. We found as much as 349 different proteins with at least one peak with signal strength >300 AU (data not shown). These 349 proteins constituted the top 20% of the genome and included spy0469, spy1643 and spy1801 as well as other antigens with published protective capacity (e.g. the M1 protein3,4, Streptolysin O42 and the streptococcal C5a peptidase33 (Fig. 3 and supplementary figure 2). This suggests that the peptide array technology can be used in future screenings for protective GAS epitopes, as it is a powerful tool to identify immunogenic antigens.
It should be noted however, that it is not fully known whether a selection strategy, based entirely on antigens/epitopes that are recognized during an infection, is the best strategy to discover protective antigens. Indeed, in several cases highly recognized epitopes (both T and B cell epitopes) may not be protective43,44,45,46,47and it can be speculated that certain key antigens/epitopes have evolved to be weakly immunogenic (“cryptic”) in order to evade protective immune responses against them. In line with this, the highly protective hypervariable region of the M protein is weakly immunogenic in infected mice and humans, but immunization with the M protein in a strong adjuvant that can overcome the low immunogenicity results in protective immunity13. Interestingly, we found that antibody responses in humans against spy1228 were substantially lower than observed for spy0469 and spy1801. However, spy1228 showed the highest protection in immunized mice, indicating that high antibody titers in humans may not always be the best predictor of protective antigens, something that has also been showed for other infections43,44,45,46,47. In fact, inducing Ab response to cryptic epitopes has for viral pathogens been shown to result in increased protection48,49, and future Streptococcus antigen discovery strategies may consider this.
In summary, we have shown that the immune response against GAS includes both antibody and cellular responses against numerous different non-M protein antigens. Using immune recognition in humans as a selection criterion for evaluation in a murine skin infection model, we identified three protective non M-protein antigens.
Methods
Antigens
Full-length sequences of antigens upregulated in four microarray studies28,29,30,31 were obtained from Streptococcus pyogenes M1 SF370. Conservation of these antigens was evaluated by a BLAST search in a local database consisting of the 26 fully sequenced GAS genomes that were available at the time of the study (see Supplementary table 1). The genomes in the database were downloaded from the NCBI genome database (http://www.ncbi.nlm.nih.gov/genome/genomes/). Based on either being predicted to have an extracellular location with the pSORT v3.0 online software or to be integral to the membrane with extracellular domains using the TMpred server32, 21 conserved antigens were selected. Sequences of the 21 antigens were codon-optimized for expression in Escherichia coli (E. coli). Transmembrane helices predicted by the TMpred server, were deleted for ease of expression and remaining fragments were linked by a sequence of -(GGGGS)2– (see Supplementary table 2 for detailed information on individual antigens). The sequences were made by chemical synthesis with an N-terminal His6-Tag followed by insertion into the commercial expression vector pJexpress 411 (DNA2.0). After transformation of E. coli BL21 AI (Invitrogen), protein expression was induced with 1 mM isopropyl β-D-1-thiogalactopyranoside (IPTG) and recombinant protein was purified as previously described50. Heat inactivated GAS bacteria (HGAS) were made by harvesting GAS colonies from blood agar plates in Tris-HCL buffer pH 7.5 before determining the bacterial concentration by plating. Bacteria were then diluted to 109 CFU/ml and killed by heating the suspension for 120 min at 60 °C. As positive controls in western blotting experiments with human heart tissue samples, we also used recombinant streptococcal M proteins from M1 (SF370) and M5 (Manfredo), which were purified in the same way as described above.
The recombinant proteins selected for protection experiments were verified by Mass spectrometry after purification (data not shown).
Human subjects
We enrolled two groups of volunteers; 32 healthy adults aged >20 years, and 30 school-aged children from 5–15 years as part of another recent study21. A simple questionnaire was used to screen the donors and all of them replied that they had not had a “sore throat” for at least 3 months before the blood samples were taken. The study was carried out in accordance with the regulations set forward by the Danish Ministry of Health and approved by the Committee on Health Research Ethics in the Capital Region (protocol no. H-2-2014-057) and the Danish Data Protection Agency (J. no. 2014-54-0733). Informed consent was obtained from all subjects.
Lymphocyte cultures
PBMCs were purified from fresh heparinized blood using a density gradient. After washing, cells were incubated at 37 °C in round bottom 96-well microtiter plates (Nunc) in 200 μl serum-free AIM-V medium (Gibco; Invitrogen) containing 3 × 105 human cells, or 2 × 105 murine cells in 200 μl RPMI-1640 supplemented with 5 × 10−5 M 2-mercaptoethanol, 1 mM glutamine, 1% pyruvate, 1% penicillin-streptomycin, 1% HEPES and 10% fetal calf serum (FCS) (Gibco; Invitrogen). Recombinant antigens were used in concentrations of 5 μg/ml and as a positive control streptococcal Enterotoxin B was used in 1 μg/ml. Supernatants in duplicates or triplicates were harvested from cultures after 3–7 days for IFNγ ELISA.
IFNγ ELISA
A sandwich ELISA was used to determine the concentration of IFNγ in culture supernatants as previously described for human samples21 and murine samples51.
Detection of antigen-specific antibodies by ELISA
Maxisorp micro titer plates (Nunc, Maxisorp) were coated with the individual recombinant antigens in a concentration of 0.5 μg/ml and antibody levels in serum/plasma samples were determined by ELISA as previously described21. In brief, free binding sites were blocked with 3% skimmed milk (w/v) and plates were incubated with individual plasma samples in 10-fold serial dilutions starting with a 1:10 dilution. Antigen-specific IgG was detected with HRP-conjugated secondary antibodies which for human samples was polyclonal rabbit anti-human IgG (Dako) diluted 1:6000 or rabbit anti-mouse IgG (Zymed) diluted 1∶5000 for murine samples. Substrate was TMB-PLUS (Kem-En-TEC). Reciprocal serum dilutions corresponding to 50% maximal binding (EC50) were computed using the GraphPad Prism 6.04.
SDS-PAGE and Western blot analysis
4–20% Mini-PROTEAN® TGX precast gels (Bio-Rad, Hercules, CA) were used, and 20 μg of human heart extract (sc-363763 from Santa Cruz Biotechnology, Dallas, TX) or human heart aorta valve whole tissue lysate (NB820-59218 from Novus Biologicals, Littleton, CO) were applied in each lane. Proteins were transferred to nitrocellulose by the Trans-Blot® Turbo™ transfer system (BioRad), the nitrocellulose was blocked in PBS with 5% skimmed milk, 0.1% Tween 20 and thereafter incubated with mouse serum samples diluted 1:200. Antigen-specific immunoglobulins were detected with alkaline phosphatase conjugated secondary antibodies.
High-density peptide arrays of selected antigens
A peptide array spanning the entire length of all predicted proteins in the entire genome of the M1 SF370 GAS strain (1,696 predicted proteins) was synthesized with 15-mers overlapping with 14 amino acids (~503,000 individual peptide fields). The layout of the array was made with proprietary software using the FASTA sequence of GAS M1 SF370 as input. Peptides spanning human serum albumin, were also included on the array as a negative control protein that does not bind human IgG. All peptides were randomly distributed on the chip to reduce the possible effect of local area-specific noise. Arrays were made by Schafer-N® using maskless photolithographic synthesis adapted to solid phase peptide synthesis with the peptide C-terminal linked to the surface of the array. The microarray was incubated for 2 hours with a 1:200 dilution of pooled plasma from six randomly selected human adults. After washing the arrays were stained for 2 hours with 1 μg/ml of a Cy3-conjugated goat anti-human detection antibody. Images of stained arrays were recorded using an InnoScan900 micro array scanner (Innopsys) with an excitation wavelength of 532 nm and fluorescence intensity for each peptide field was calculated by summarizing over the R, G and B channels in the resulting image file.
Bacterial strains and growth
GAS strain MGAS5005 (serotype M1) was grown at 37 °C with 5% CO2 in Todd-Hewitt broth (SSI Diagnostica) or on 5% blood agar (SSI Diagnostica) that was used as solid medium.
Animal experiments
Female DBA/2 mice at 5–8 weeks of age were purchased at Harlan Laboratories (Horst, The Netherlands) and randomized to cages upon arrival. Handling of the animals was conducted in accordance with the regulations of the Danish Ministry of Justice and animal protection committees and in compliance with the EU Directive 2010/63/EU. The methods were carried out in accordance with the approved guidelines. All animal experiments were approved by an institutional animal committee. Mice were immunized subcutaneously (s.c.) at the base of the tail with 5 μg of recombinant antigen formulated in AddaVaxTM 1:1 (Invivogen). Three immunizations were performed with a two-week interval and four weeks after last immunization, mice received an intradermal (i.d.) injection of 0.5–1 × 107 CFU (GAS strain MGAS M5005, M1 serotype). The mice were sacrificed 4 days post infection and skin biopsies were obtained by excising a specimen of 1–1.5 cm in diameter around the infected area. Vaccine-induced protection was evaluated by counting the CFUs obtained from plating suitable dilutions of skin homogenates.
Statistics
CFU levels in challenged mice were compared by a student’s t-test in two independent animal experiments for each antigen. A chi-square test was used to compare responder frequencies in human adults and children. p < 0.05 was considered significant. All statistical analyses were carried out in GraphPad Prism version 6.04 (GraphPad Software Inc.).
Additional Information
How to cite this article: Mortensen, R. et al.Identifying protective Streptococcus pyogenesvaccine antigens recognized by both B and T cells in human adults and children. Sci. Rep. 6, 22030; doi: 10.1038/srep22030 (2016).
References
- 1.
Cunningham, M. W. Pathogenesis of group A streptococcal infections. Clin Microbiol Rev 13, 470–511 (2000).
- 2.
Carapetis, J. R., Steer, A. C., Mulholland, E. K. & Weber, M. The global burden of group A streptococcal diseases. Lancet Infect Dis5, 685–694 (2005).
- 3.
Dale, J. B., Penfound, T. A., Chiang, E. Y. & Walton, W. J. New 30-valent M protein-based vaccine evokes cross-opsonic antibodies against non-vaccine serotypes of group A streptococci. Vaccine 29, 8175–8178 (2011).
- 4.
Caro-Aguilar, I. et al. Immunogenicity in mice and non-human primates of the Group A Streptococcal J8 peptide vaccine candidate conjugated to CRM197. Hum Vaccin Immunother 9, 488–496 (2013).
- 5.
Zaman, M. et al. Immunological evaluation of lipopeptide group A streptococcus (GAS) vaccine: structure-activity relationship. PLoS One 7, e30146 (2012).
- 6.
Postol, E. et al. StreptInCor: a candidate vaccine epitope against S. pyogenes infections induces protection in outbred mice. PLoS One 8, e60969 (2013).
- 7.
Sanderson-Smith, M. et al. A systematic and functional classification of Streptococcus pyogenes that serves as a new tool for molecular typing and vaccine development. J Infect Dis 210, 1325–1338, (2014).
- 8.
Fae, K. C. et al. Mimicry in recognition of cardiac myosin peptides by heart-intralesional T cell clones from rheumatic heart disease. J Immunol 176, 5662–5670 (2006).
- 9.
Martins, T. B. et al. Comprehensive analysis of antibody responses to streptococcal and tissue antigens in patients with acute rheumatic fever. Int immunol 20, 445–452 (2008).
- 10.
Steer, A. C., Batzloff, M. R., Mulholland, K. & Carapetis, J. R. Group A streptococcal vaccines: facts versus fantasy. Curr Opin Infect Dis 22, 544–552 (2009).
- 11.
Steer, A. C., Dale, J. B. & Carapetis, J. R.Progress toward a global group a streptococcal vaccine. Pediatr Infect Dis J32, 180–182 (2013).
- 12.
Bessen, D. & Fischetti, V. A. Passive acquired mucosal immunity to group A streptococci by secretory immunoglobulin A. J Exp Med167, 1945–1950 (1988).
- 13.
Lannergard, J. et al. The Hypervariable region of Streptococcus pyogenes M protein escapes antibody attack by antigenic variation and weak immunogenicity. Cell Host Microbe 10, 147–157 (2011).
- 14.
Pandey, M., Batzloff, M. R. & Good, M. F.Mechanism of protection induced by group A Streptococcus vaccine candidate J8-DT: contribution of B and T-cells towards protection. PLoS One 4, e5147 (2009).
- 15.
Dale, J. B., Chiang, E. Y., Liu, S., Courtney, H. S. & Hasty, D. L. New protective antigen of group A streptococci. J Clin Invest 103, 1261–1268 (1999).
- 16.
Park, H. S. & Cleary, P. P. Active and passive intranasal immunizations with streptococcal surface protein C5a peptidase prevent infection of murine nasal mucosa-associated lymphoid tissue, a functional homologue of human tonsils. Infect Immun 73, 7878–7886 (2005).
- 17.
Siegert, J., Sastalla, I., Chhatwal, G. S. & Medina, E. Vaccination equally enables both genetically susceptible and resistant mice to control infection with group A streptococci. Microbes Infect 8, 347–353 (2006).
- 18.
Fan, X. et al. Sortase a induces th17-mediated and antibody-independent immunity to heterologous serotypes of group a streptococci. PLoS One 9, e107638 (2014).
- 19.
Wang, B. et al. Induction of TGF-beta1 and TGF-beta1-dependent predominant Th17 differentiation by group A streptococcal infection. Proc Natl Acad Sci USA 107, 5937–5942 (2010).
- 20.
Dileepan, T. et al. Robust antigen specific th17 T cell response to group A Streptococcus is dependent on IL-6 and intranasal route of infection. PLoS Pathog 7, e1002252 (2011).
- 21.
Mortensen, R. et al. Adaptive Immunity against Streptococcus pyogenes in Adults Involves Increased IFN-gamma and IgG3 Responses Compared with Children. J Immunol 195, 1657–64 (2015).
- 22.
Lei, B., Liu, M., Chesney, G. L. & Musser, J. M. Identification of new candidate vaccine antigens made by Streptococcus pyogenes: purification and characterization of 16 putative extracellular lipoproteins. J Infect Dis 189, 79–89 (2004).
- 23.
Rodriguez-Ortega, M. J. et al.Characterization and identification of vaccine candidate proteins through analysis of the group A Streptococcus surface proteome. Nat biotechnol 24, 191–197 (2006).
- 24.
Fritzer, A. et al. Novel conserved group A streptococcal proteins identified by the antigenome technology as vaccine candidates for a non-M protein-based vaccine. Infect Immun 78, 4051–4067 (2010).
- 25.
Bensi, G. et al. Multi high-throughput approach for highly selective identification of vaccine candidates: the Group A Streptococcus case. Mol Cell Proteomics 11, M111 015693, (2012).
- 26.
Henningham, A. et al. Conserved anchorless surface proteins as group A streptococcal vaccine candidates. J Mol Med (Berl) 90, 1197–1207, (2012).
- 27.
McMillan, D. J. et al. Identification and assessment of new vaccine candidates for group A streptococcal infections. Vaccine22, 2783–2790 (2004).
- 28.
Virtaneva, K. et al. Longitudinal analysis of the group A Streptococcus transcriptome in experimental pharyngitis in cynomolgus macaques. Proc Natl Acad Sci USA 102, 9014–9019 (2005).
- 29.
Graham, M. R. et al. Group A Streptococcus transcriptome dynamics during growth in human blood reveals bacterial adaptive and survival strategies. Am J Pathol 166, 455–465 (2005).
- 30.
Graham, M. R. et al. Analysis of the transcriptome of group A Streptococcus in mouse soft tissue infection. Am J Pathol 169, 927–942 (2006).
- 31.
Voyich, J. M. et al. Genome-wide protective response used by group A Streptococcus to evade destruction by human polymorphonuclear leukocytes. Proc Natl Acad Sci USA 100, 1996–2001 (2003).
- 32.
Hofmann, K. & Stoffel, W. TMbase-A database of membrane spanning protein segments. Biol Chem 374, 166 (1993).
- 33.
Ji, Y., Carlson, B., Kondagunta, A. & Cleary, P. P. Intranasal immunization with C5a peptidase prevents nasopharyngeal colonization of mice by the group A Streptococcus. Infect Immun 65, 2080–2087 (1997).
- 34.
Tsai, L. M. & Yu, D. Follicular helper T-cell memory: establishing new frontiers during antibody response. Immunol Cell Biol 92, 57–63 (2014).
- 35.
Sutcliffe, I. C. & Harrington, D. J. Pattern searches for the identification of putative lipoprotein genes in Gram-positive bacterial genomes. Microbiology 148, 2065–2077 (2002).
- 36.
Okamoto, A., Hasegawa, T., Yamada, K. & Ohta, M. Application of both high-performance liquid chromatography combined with tandem mass spectrometry shotgun and 2-D polyacrylamide gel electrophoresis for streptococcal exoproteins gave reliable proteomic data. Microbiol Immunol 55, 84–94 (2011).
- 37.
Buist, G., Steen, A., Kok, J. & Kuipers, O. P.LysM, a widely distributed protein motif for binding to (peptido)glycans. Mol Microbiol68, 838–847 (2008).
- 38.
Li, J. et al. Neutrophils select hypervirulent CovRS mutants of M1T1 group A Streptococcus during subcutaneous infection of mice. Infect Immun 82, 1579–1590 (2014).
- 39.
Cho, K. H. & Caparon, M. G. Patterns of virulence gene expression differ between biofilm and tissue communities of Streptococcus pyogenes. Mol Microbiol 57, 1545–1556 (2005).
- 40.
Mercier, C. et al. Positive role of peptidoglycan breaks in lactococcal biofilm formation. Mol Microbiol 46, 235–243 (2002).
- 41.
McIver, K. S., Subbarao, S., Kellner, E. M., Heath, A. S. & Scott, J. R. Identification of isp, a locus encoding an immunogenic secreted protein conserved among group A streptococci. Infect Immun 64, 2548–2555 (1996).
- 42.
Chiarot, E. et al. Targeted amino acid substitutions impair streptolysin O toxicity and group A Streptococcus virulence. MBio4, e00387–00312 (2013).
- 43.
Nara, P. L. & Garrity, R. Deceptive imprinting: a cosmopolitan strategy for complicating vaccination. Vaccine 16, 1780–1787 (1998).
- 44.
Kohler, H., Goudsmit, J. & Nara, P. Clonal Dominance-Cause for a Limited and Failing Immune-Response to Hiv-1 Infection and Vaccination. J Acq Immun Def Synd 5, 1158–1168 (1992).
- 45.
Sugimoto, K. et al. Suppression of HCV-specific T cells without differential hierarchy demonstrated ex vivo in persistent HCV infection. Hepatology 38, 1437–1448 (2003).
- 46.
Friedrich, T. C. et al. Subdominant CD8(+) T-cell responses are involved in durable control of AIDS virus replication. J Virol 81, 3465–3476 (2007).
- 47.
Aagaard, C. S., Hoang, T. T. K. T., Vingsbo-Lundberg, C., Dietrich, J. & Andersen, P.Quality and Vaccine Efficacy of CD4(+) T Cell Responses Directed to Dominant and Subdominant Epitopes in ESAT-6 from Mycobacterium tuberculosis. J Immunol183, 2659–2668 (2009).
- 48.
Garrity, R. R. et al. Refocusing neutralizing antibody response by targeted dampening of an immunodominant epitope. J Immunol159, 279–289 (1997).
- 49.
Tobin, G. J. et al. Deceptive imprinting and immune refocusing in vaccine design. Vaccine 26, 6189–6199 (2008).
- 50.
Aagaard, C. et al. A multistage tuberculosis vaccine that confers efficient protection before and after exposure. Nat Med 17, 189–194 (2011).
- 51.
Lindenstrom, T. et al. Vaccine-induced th17 cells are maintained long-term postvaccination as a distinct and phenotypically stable memory subset. Infect Immun 80, 3533–3544 (2012).
Acknowledgements
The authors thank the volunteers participating in the study and gratefully acknowledge Lene Rasmussen, Vivi Andersen and Janne Rabech for excellent technical assistance at Statens Serum Institut. This study was supported by University of Copenhagen, Familien Hede Nielsens Fond, A.P. Møller Fonden, The Novo Nordisk Foundation, Brd. Hartmann Fonden, Beckett Fonden and Centre for Nano-vaccines, Grant # 0603-00322B. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Supplementary information
PDF files
Rights and permissions
 This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/